
I have been studyin up. I ran across this, most of you probablly already know about. If it can help simplfy breeding then its well worth it.
MENDEL's work is outstanding, since it constitutes a completely new approach: the concentration on just a single feature. MENDEL set great store by the evaluation of the numerical proportions of the hybrids and he analyzed the plants gained by hybridization independently. He found it also essential to work with as great a number as plants as possible in order to outrule chance. His research enabled him to detect three principles of heredity.
MENDEL's first law is the principle of uniformity. It says that, if two plants that differ in just one trait are crossed, then the resulting hybrids will be uniform in the chosen trait. Depending on the traits is the uniform feature either one of the parents' traits (a dominant-recessive pair of characteristics) or it is intermediate.
MENDEL's second law is the principle of segregation. It states that the individuals of the F2 generation are not uniform, but that the traits segregate. Depending on a dominant-recessive crossing or an intermediate crossing are the resulting ratios 3:1 or 1:2:1. According to this principle hereditary traits are determined by discrete factors (now called genes) that occur in pairs, one of each pair being inherited from each parent. This concept of independent traits explains how a trait can persist from generation to generation without blending with other traits. It explains, too, how the trait can seemingly disappear and then reappear in a later generation. The principle of segregation was consequently of the utmost importance for understanding both genetics and evolution.
MENDEL's third law is also called the principle of independent assortment. It says that every trait is inherited independently of the others and it thus covers the case that new combinations of genes can arise, which were not existing before. We know today that this principle is just valid in the case of genes that are not coupled, i.e. that are not located at the same chromosome.
in 1866 that was to become the foundation of modern genetics. In contrast to his predecessors MENDEL was not interested in the problem of the sexuality of flowering plants and not in the delimitation of species and varieties, his interest was the numerical registration of the transmission of parental properties to the hybrids. He approached it in a completely new way that differed from those used before in three points:
First, he did not hybridize species and varieties that differed in many properties. In contrast, the first big success of MENDEL's method was the isolated examination of a single feature. Second, he attached great importance to the numerical proportions of the hybrids. He wrote:
"It seemed necessary that all members of a developmental line were completely examined in each respective generation in order to recognize the relations, that the different types of hybrids have to each other and to their parental species."
MENDEL found it essential to work with as big a number of individuals as possible, because chance could otherwise exert too great an influence and thus conceal the rules that were visible at appropriately large numbers. Third, he analyzed the plants he gained by hybridization independently. In the same careful manner he did also separate the respective generations of bastards. He wrote about his plants:
"The choice of the plant groups has to occur with outmost care, if not the whole success has to be questioned. It is necessary that the plants have
1. constantly differing traits.
2. Their hybrids have to be protected from the influence of any foreign pollen during flowering or it has to be easy to protect them upon need.
3. The hybrids and their offspring are not to suffer from any impairment of fertility.
He chose Pisum sativum as his specimen and concentrated on the analysis of seven pairs of characteristics. The choice of the object was happy, since the garden pea is generally self-pollinating. Accordingly the varieties he used were true-breeding. MENDEL concluded the following:
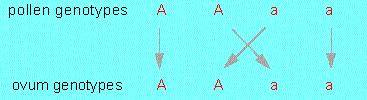
If two plants that differ from each other in just one characteristic are crossed, then the resulting hybrids are uniform in the chosen characteristic. This statement is today known as the MENDEL's first law and is also called the principle of uniformity.
Before we advance to further laws, we have to explain several terms. The parental generation is marked with a P. The first daughter generation is named F1 from Latin 'filia', which means 'daughter'. If two hybrids of the F1 generation are crossed, then the resulting generation is called F2, the following generation F3 and so on. One of the two characteristics of a pair may be (as actually all pairs chosen by MENDEL were) dominant. Such a crossing is also called dominant-recessive. All individuals of the F1 generation bear the dominant characteristic. Contrasting are crossings where the resulting characteristic of the F1 generation is an intermediate of the two parental ones. Such a crossing is called intermediate or incompletely dominant.
For both types of crossing it is unimportant, whether the characteristic stems from the male or the female parent. An example shall elucidate this: (female) round seeds x (male) edgy seeds result in F1 individuals with round seeds , since round is dominant. The same result is achieved with the following cross: (female) edgy seeds x (male) round seeds. In a cross is the first mentioned parent always the female one.
If now the hybrids are crossed with each other, then the recessive characteristic occurs again in the F2 generation (see table). MENDEL extrapolated a ratio of 3:1 from his experimental results. He concluded:
"Since the members of the first generation of hybrids (meant is the F2 generation) stem directly from the seeds of the hybrids, it now becomes clear that the hybrids of two different characteristics form seeds, of which one half develops again into the hybrid state, while the other half results in plants, which stay constant in an either dominant or recessive character. Dominant and recessive characteristic do show in equal numbers here."
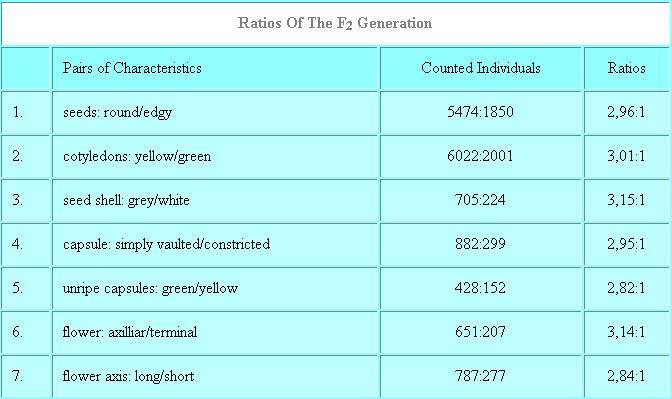
MENDEL assumed that both pollen and eggs bear hereditary factors, that he thought to be something different than the characteristics themselves. He made thus a further important insight, because not the characteristics themselves are inherited, but their lay-out (now called alleles).
Since it is dependent on chance, which pollen and eggs combine, the pollination will take place according to the rules of probability as shown in the following scheme:
This results in the offspring:
AA Aa aA aa
This regularity is expressed by MENDEL's second law, the principle of segregation. The individuals of the F2 generation are not uniform, instead different types are visible. The characteristics of the parental generation do always occur at a certain ratio. Depending on a dominant-recessive or an intermediate crossing, they segregate in the ratio 3:1 or 1:2:1.
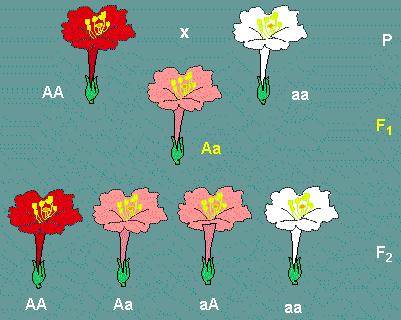
Incomplete dominance in flowers of Mirabilis jalapa AA genotypes have red, Aa genotypes pink and aa genotypes whitish flowers (redrawn from C. CORRENS, 1902)
It cannot be perceived whether the dominant individuals of a dominant-recessive hereditary path will breed pure or whether the traits will segregate again in following generations. A formal reflection will explain this. A hereditary factor like, for example, the shape of the seed, the colour of the cotyledons or the colour of the seed shell shall be called a gene (following a suggestion of BATESON made in 1909). The state, in which a gene exists, i.e. whether the trait 'seed shape' is, for example, round or edgy is called an allele. If we look at the scheme above, we will see that each individual inherits one allele per gene of the mother and one of the father. Both alleles may be equal or they may differ. In the first case the individual would be homozygous, in the latter heterozygous. MENDEL did already mark genes (which he called 'Anlagen' (German), meaning 'hereditary factors') by letters (see above). Here, too, international conventions do nowadays exist that fix the way, in which these letters are written. Nevertheless different terminologies are common in the different disciplines of genetics. Drosophila researchers , for example, use other terms than bacterial researchers.
But let us start with a fundamental rule. A dominant allele is marked with a capital, a recessive one with the respective lowercase letter. So if a plant with red flowers is crossed with a plant with white flowers and red is dominant (white is accordingly recessive), it cannot be operated with the letters r (for red) and w (for white), but the red flowers have to be marked R and the white ones r. Since two alleles exist for each gene in each individual, the homozygous forms RR and rr and the heterozygous Rrexist.
Homozygous individuals (RR) cannot be distinguished from heterozygous ones (Rr) in dominant-recessive hereditary paths. To be able to recognize their true type the hybrids are crossed with the recessive parent (rr). This is called a back-cross. Homozygous individuals (RR x rr) produce only one type of offspring (Rr), while the offspring of heterozygous individuals (Rr x rr) is half Rr and half rr.
Since it cannot easily be distinguished between a homozygous and a heterozygous state (RR or Rr), it is instead differentiated between phenotype (appearance) and genotype (the allele combination). A certain phenotype can thus have several (two in the case of a dominant phenotype in a hereditary path of two alleles per gene) genotypes.
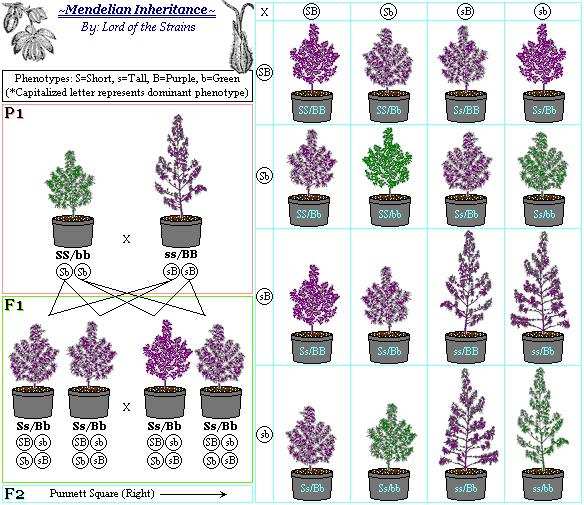
Now, what happens, if several pairs of characteristics are regarded? MENDEL's example was the following: round seeds and yellow cotyledons x edgy seeds and green cotyledons. Both plants were homozygous for the respective characters.
All hybrids of the F1 generation had round seeds and yellow cotyledons, but in the F2 generation a segregation could be observed:
315 plants had round seeds and yellow cotyledons,
101 had edgy seeds and yellow cotyledons,
38 inherited edgy seeds and green cotyledons and
108 plants had round seeds and green cotyledons
This result could be interpreted by the following scheme:
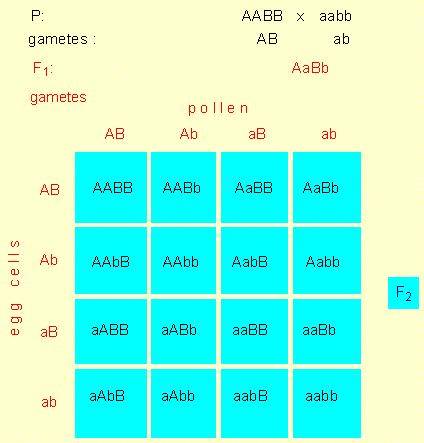
PUNNETT-Square: The scheme shows the genotypes of the P-, F1- and F2-generation of a dihybrid hereditary path. This kind of representation was introduced by the British geneticist R. C. PUNNETT at the beginning of this century
The experiment proved that the characters were inherited independently from one another. The genotypes of the F2 generation occur in a ratio of 9:3:3:1. And this leads us to MENDEL's third law, the principle of independent assortment. It does inevitably cover the case that new combinations of genes, that were not existing before can arise. In MENDEL's experiment these are the combinations: round seeds/green cotyledons and edgy seeds/yellow cotyledons.
If just one character is studied, then it is talked of a monohybrid crossing. If further characteristics are also regarded, then the crossing is called dihybrid, trihybrid,...polyhybrid.
this pic should be in there somewhere I lost its place!
MENDEL's work is outstanding, since it constitutes a completely new approach: the concentration on just a single feature. MENDEL set great store by the evaluation of the numerical proportions of the hybrids and he analyzed the plants gained by hybridization independently. He found it also essential to work with as great a number as plants as possible in order to outrule chance. His research enabled him to detect three principles of heredity.
MENDEL's first law is the principle of uniformity. It says that, if two plants that differ in just one trait are crossed, then the resulting hybrids will be uniform in the chosen trait. Depending on the traits is the uniform feature either one of the parents' traits (a dominant-recessive pair of characteristics) or it is intermediate.
MENDEL's second law is the principle of segregation. It states that the individuals of the F2 generation are not uniform, but that the traits segregate. Depending on a dominant-recessive crossing or an intermediate crossing are the resulting ratios 3:1 or 1:2:1. According to this principle hereditary traits are determined by discrete factors (now called genes) that occur in pairs, one of each pair being inherited from each parent. This concept of independent traits explains how a trait can persist from generation to generation without blending with other traits. It explains, too, how the trait can seemingly disappear and then reappear in a later generation. The principle of segregation was consequently of the utmost importance for understanding both genetics and evolution.
MENDEL's third law is also called the principle of independent assortment. It says that every trait is inherited independently of the others and it thus covers the case that new combinations of genes can arise, which were not existing before. We know today that this principle is just valid in the case of genes that are not coupled, i.e. that are not located at the same chromosome.
in 1866 that was to become the foundation of modern genetics. In contrast to his predecessors MENDEL was not interested in the problem of the sexuality of flowering plants and not in the delimitation of species and varieties, his interest was the numerical registration of the transmission of parental properties to the hybrids. He approached it in a completely new way that differed from those used before in three points:
First, he did not hybridize species and varieties that differed in many properties. In contrast, the first big success of MENDEL's method was the isolated examination of a single feature. Second, he attached great importance to the numerical proportions of the hybrids. He wrote:
"It seemed necessary that all members of a developmental line were completely examined in each respective generation in order to recognize the relations, that the different types of hybrids have to each other and to their parental species."
MENDEL found it essential to work with as big a number of individuals as possible, because chance could otherwise exert too great an influence and thus conceal the rules that were visible at appropriately large numbers. Third, he analyzed the plants he gained by hybridization independently. In the same careful manner he did also separate the respective generations of bastards. He wrote about his plants:
"The choice of the plant groups has to occur with outmost care, if not the whole success has to be questioned. It is necessary that the plants have
1. constantly differing traits.
2. Their hybrids have to be protected from the influence of any foreign pollen during flowering or it has to be easy to protect them upon need.
3. The hybrids and their offspring are not to suffer from any impairment of fertility.
He chose Pisum sativum as his specimen and concentrated on the analysis of seven pairs of characteristics. The choice of the object was happy, since the garden pea is generally self-pollinating. Accordingly the varieties he used were true-breeding. MENDEL concluded the following:
If two plants that differ from each other in just one characteristic are crossed, then the resulting hybrids are uniform in the chosen characteristic. This statement is today known as the MENDEL's first law and is also called the principle of uniformity.
Before we advance to further laws, we have to explain several terms. The parental generation is marked with a P. The first daughter generation is named F1 from Latin 'filia', which means 'daughter'. If two hybrids of the F1 generation are crossed, then the resulting generation is called F2, the following generation F3 and so on. One of the two characteristics of a pair may be (as actually all pairs chosen by MENDEL were) dominant. Such a crossing is also called dominant-recessive. All individuals of the F1 generation bear the dominant characteristic. Contrasting are crossings where the resulting characteristic of the F1 generation is an intermediate of the two parental ones. Such a crossing is called intermediate or incompletely dominant.
For both types of crossing it is unimportant, whether the characteristic stems from the male or the female parent. An example shall elucidate this: (female) round seeds x (male) edgy seeds result in F1 individuals with round seeds , since round is dominant. The same result is achieved with the following cross: (female) edgy seeds x (male) round seeds. In a cross is the first mentioned parent always the female one.
If now the hybrids are crossed with each other, then the recessive characteristic occurs again in the F2 generation (see table). MENDEL extrapolated a ratio of 3:1 from his experimental results. He concluded:
"Since the members of the first generation of hybrids (meant is the F2 generation) stem directly from the seeds of the hybrids, it now becomes clear that the hybrids of two different characteristics form seeds, of which one half develops again into the hybrid state, while the other half results in plants, which stay constant in an either dominant or recessive character. Dominant and recessive characteristic do show in equal numbers here."
MENDEL assumed that both pollen and eggs bear hereditary factors, that he thought to be something different than the characteristics themselves. He made thus a further important insight, because not the characteristics themselves are inherited, but their lay-out (now called alleles).
Since it is dependent on chance, which pollen and eggs combine, the pollination will take place according to the rules of probability as shown in the following scheme:
This results in the offspring:
AA Aa aA aa
This regularity is expressed by MENDEL's second law, the principle of segregation. The individuals of the F2 generation are not uniform, instead different types are visible. The characteristics of the parental generation do always occur at a certain ratio. Depending on a dominant-recessive or an intermediate crossing, they segregate in the ratio 3:1 or 1:2:1.
Incomplete dominance in flowers of Mirabilis jalapa AA genotypes have red, Aa genotypes pink and aa genotypes whitish flowers (redrawn from C. CORRENS, 1902)
It cannot be perceived whether the dominant individuals of a dominant-recessive hereditary path will breed pure or whether the traits will segregate again in following generations. A formal reflection will explain this. A hereditary factor like, for example, the shape of the seed, the colour of the cotyledons or the colour of the seed shell shall be called a gene (following a suggestion of BATESON made in 1909). The state, in which a gene exists, i.e. whether the trait 'seed shape' is, for example, round or edgy is called an allele. If we look at the scheme above, we will see that each individual inherits one allele per gene of the mother and one of the father. Both alleles may be equal or they may differ. In the first case the individual would be homozygous, in the latter heterozygous. MENDEL did already mark genes (which he called 'Anlagen' (German), meaning 'hereditary factors') by letters (see above). Here, too, international conventions do nowadays exist that fix the way, in which these letters are written. Nevertheless different terminologies are common in the different disciplines of genetics. Drosophila researchers , for example, use other terms than bacterial researchers.
But let us start with a fundamental rule. A dominant allele is marked with a capital, a recessive one with the respective lowercase letter. So if a plant with red flowers is crossed with a plant with white flowers and red is dominant (white is accordingly recessive), it cannot be operated with the letters r (for red) and w (for white), but the red flowers have to be marked R and the white ones r. Since two alleles exist for each gene in each individual, the homozygous forms RR and rr and the heterozygous Rrexist.
Homozygous individuals (RR) cannot be distinguished from heterozygous ones (Rr) in dominant-recessive hereditary paths. To be able to recognize their true type the hybrids are crossed with the recessive parent (rr). This is called a back-cross. Homozygous individuals (RR x rr) produce only one type of offspring (Rr), while the offspring of heterozygous individuals (Rr x rr) is half Rr and half rr.
Since it cannot easily be distinguished between a homozygous and a heterozygous state (RR or Rr), it is instead differentiated between phenotype (appearance) and genotype (the allele combination). A certain phenotype can thus have several (two in the case of a dominant phenotype in a hereditary path of two alleles per gene) genotypes.
Now, what happens, if several pairs of characteristics are regarded? MENDEL's example was the following: round seeds and yellow cotyledons x edgy seeds and green cotyledons. Both plants were homozygous for the respective characters.
All hybrids of the F1 generation had round seeds and yellow cotyledons, but in the F2 generation a segregation could be observed:
315 plants had round seeds and yellow cotyledons,
101 had edgy seeds and yellow cotyledons,
38 inherited edgy seeds and green cotyledons and
108 plants had round seeds and green cotyledons
This result could be interpreted by the following scheme:
PUNNETT-Square: The scheme shows the genotypes of the P-, F1- and F2-generation of a dihybrid hereditary path. This kind of representation was introduced by the British geneticist R. C. PUNNETT at the beginning of this century
The experiment proved that the characters were inherited independently from one another. The genotypes of the F2 generation occur in a ratio of 9:3:3:1. And this leads us to MENDEL's third law, the principle of independent assortment. It does inevitably cover the case that new combinations of genes, that were not existing before can arise. In MENDEL's experiment these are the combinations: round seeds/green cotyledons and edgy seeds/yellow cotyledons.
If just one character is studied, then it is talked of a monohybrid crossing. If further characteristics are also regarded, then the crossing is called dihybrid, trihybrid,...polyhybrid.
this pic should be in there somewhere I lost its place!
Last edited:


") nice work GPB
nice work GPB 

 Thanks for lettin me vent, that still makes me mad.
Thanks for lettin me vent, that still makes me mad.
