
")
denke mal das einige schon davon gehört haben is ja auch schon paar jährchen alt der artikel,
aber vielleicht reichts bei euch wie bei mir auch zu ner zehnminütigen unterhaltung der synapsen....
hier findet ihr die originalstory1Killing Cannabis with mycoherbicides
John M. McPartland
VAM/AMRITA, 53 Washington Street Extension, Middlebury, VT 05753, USA
e-mail: [email protected], phone: 802-388-0575, fax: 802-382-8845
David West
GamETec, 363 S. Warren Street, Prescott, WI 54021, USA
McPartland, John M. and David West 1999. Killing Cannabis with mycoherbicides Journal of the International Hemp Association 6(1): 1, 4-8. Last year, researchers were funded by the U.S. government to create fungi that destroy drug plants, including marijuana (Cannabis). The fungi will be genetically engineered. Controversies surrounding this "new solution" for the war on drugs are discussed, including the ethics of exterminating plant species that have occupied central roles in human culture for thousands of years. The importation of foreign fungi into new habitats is fraught with unpredictable environmental pitfalls; exotic pathogens can spread from their intended targets to other organisms. All known pathogens of marijuana also attack hemp; exterminating drug plants will probably spell the demise of the valuable and resurgent fiber and oil-seed crop. Genetically transformed fungi are genetically unstable and mutate easily. Fungi with recombinant DNA may reproduce with native fungi and create new strains of virulent, transgenic pathogens. Once these pathogens are released in the environment, they cannot be recalled. In summary, research involving transgenic pathogens of Cannabis is a dangerous misuse of biotechnology, and should be the subject of an immediate moratorium.
 IntroductionFigure 1. Healthy marijuana seedling (C) flanked by plants exposed to pathogenic fungi (P.g. and M.p.).
IntroductionFigure 1. Healthy marijuana seedling (C) flanked by plants exposed to pathogenic fungi (P.g. and M.p.).
The U.S. Congress recently appropriated $23 million dollars to fund a "new solution" for the war on drugs. The new solution attacks drugs at their source — the drug plants. Researchers say they can eliminate drug plants with fungal pathogens. The fungi would be genetically engineered to kill only coca plants (Erythroxylon sp.), opium poppies (Papaver sp.), and marijuana (Cannabis sp.).
Rep. Bill McCollum, who introduced the appropriation bill, described the tactic as "a silver bullet in the drug war" (Fields 1998). The development of transgenic coca and opium pathogens began several years ago, but previous appropriations were relatively small (the 1998 budget was $2.58 million). This year McCollum expanded the program to include marijuana, and moved the budget’s decimal point to the right.
A fungal weapon (Fig. 1) for the war on drugs is not new. Millions of dollars were spent in the 1970s in a world-wide search for fungi which would attack coca (Lentz et al. 1975), poppies (Schmitt and Lipscomb 1975), or marijuana (Ghani et al. 1978). It was a strange era for plant pathologists. While researchers around the globe attacked the pathogens of poppies and hemp, US-funded scientists reversed the strategy — they attacked poppies and hemp with these same pathogens (Doctor 1986).
Renewed interest in fungal pathogens for the "war on drugs" is of great concern. The law-enforcement lobby wishes to exterminate three plant species that have occupied central roles in human culture for thousands of years. Are the targeted plants inescapably evil? Are there no alternative means for reducing their dangers to humans? Reported herein are the ethical and scientific controversies pertinent to this issue, framed for consideration by academia, state and federal government agencies, and others interested in genetically engineered organisms, biological control, and the drug war (Cook et al. 1996).
Killer fungi
Experiments with fungi to control plants began in the late 1960s. The initial targets were noxious agricultural weeds that had been accidentally imported from one region of the world into another, where they became more aggressive because their natural enemies were often absent. Hence, the classical strategy for biocontrol of weeds involves the importation of natural enemies from their native ranges. Classical biocontrol generally enjoys wide approval and is used by organic agriculture, although the strategy does have its critics (Howarth 1991).
Classical biocontrol of marijuana was originally envisioned by Arthur McCain in 1970 (Shay 1975). McCain, a professor at the University of California-Berkeley, suggested, "Just introduce a couple of pounds [of a pathogenic fungus] into an area, and while it wouldn’t have much of an effect the first year, in several years it would spread throughout the country with devastating results" (Zubrin 1981). In reality, however, classical biocontrol rarely extirpates a weed, it merely reduces the weed population to a low level (Watson 1991). Reduction without eradication is acceptable for most agricultural weeds, but is unacceptable for "zero tolerance" drug control, which seeks the complete eradication of a crop.
The other biocontrol strategy, inundative release, is also called the mycoherbicide approach. This strategy releases massive amounts of fungal spores upon target plants. The mycoherbicide approach can totally eradicate a field of drug plants. This approach, however, utilizes a delivery system similar to that of chemical herbicides — such as hovering over clandestine fields in a helicopter while releasing the control agent. Thus the mycoherbicide approach, compared to the current herbicide strategy, is equally expensive, exposes pilots to equal danger as they hover over fields, and may require retreatment of annual crops. The mycoherbicide approach is not the suggested "silver bullet."
Fear of foreigners
The importation of foreign fungi into new habitats is fraught with controversy. Once a self-perpetuating fungus has been released, it is impossible to recall or control (Lockwood 1993). Despite host-range testing to identify potential nontarget hosts, exotic fungi can spread from their intended targets to other plants. The entire flora of a continent may ultimately be exposed, especially if the fungus produces wind-borne spores (Auld 1991). Because of this concern, only two exotic fungi have ever been intentionally imported into North America—Puccinia chondrillina and Puccinia carduorum.
Fear of "collateral damage" to nontarget plants is justified. When Puccinia xanthii, considered a selective pathogen of Xanthium weeds, was imported into Australia from North America, the fungus spread to sunflowers (Helianthus annuus) and Calendula officinalis (Auld 1991). Native fungi sold as mycoherbicides may also spread to new hosts after release. For example, Colletotrichum gloesporioides f. sp. aeschynomene (Collego®), one of only three mycoherbicide fungi commercially available in the U.S., has a wider host range than originally determined, including several economically important legumes (TeBeest 1988).
The situation with insects is comparable to that with fungi. Turner (1985) estimated that 21% of biocontrol insects intentionally introduced into North America have spread to non-target native plants. For instance, the beetle Chrysolina quadrigemina was imported into North America to kill weedy St. John’s wort (Hypericum perforatum), but it subsequently moved to the ornamental species Hypericum calycinum (Turner 1985). Howarth (1991) described nearly 100 cases where errant biocontrols have driven non-target hosts to extinction, mostly in island ecosystems. Howarth claimed that more species extinctions have been caused by biocontrols than by pesticides.
Non-target hosts at greatest risk to exotic biocontrol fungi include:
The study of fungus-host specificity is site-dependent. That is, each potential release site has its own unique flora, fauna, and climatic conditions. Sites with a high degree of biodiversity, such as Amazonia, are teeming with potential non-target hosts. Studies of tropical sites are very complicated and become susceptible to errors of tremendous consequence. The potential spread of fungi away from release sites must also be taken under consideration. Biocontrol agents do not recognize international boundaries, yet host specificity studies rarely consider non-target hosts in neighboring countries (Lockwood 1993).
- plants phylogenetically related to the target species,
- plants with secondary compounds or morphological features similar to the target species,
- plants attacked by fungi related to the biocontrol fungus,
- plants never exposed to the biocontrol fungus,
- plants whose fungal pathogens are unknown (Watson 1991).
In the case of pathogens of Cannabis, the non-target host at greatest risk, because of its close phylogenetic relationship to Cannabis, is hop (Humulus lupulus). At least 10 fungal pathogens are known to mutually infect Cannabis and Humulus (McPartland 1992). The next closest relatives are the Urticaceae (members of the nettle family) and the Moraceae (mulberry family), with which Cannabis shares at least 20 fungal pathogens (McPartland 1992).
The species debate
The non-target host at greatest risk is Cannabis itself. Within the genus we find plants cultivated for drugs (marijuana), or for fiber or seed (hemp), as well as feral plants. How closely related are these plants? Some taxonomists describe marijuana and hemp as completely separate species (Schultes et al. 1974), whereas other taxonomists say they are the same species, Cannabis sativa (Small and Cronquist 1976).
This "species debate" achieved semantic importance during the 1970s (Small 1979). Drug libertarians promoted the polytypic approach and cited marijuana as Cannabis indica to argue that statutes written against Cannabis sativa did not apply to marijuana. Conversely, law enforcement agencies have maintained that the genus is monotypic. Now, to rationalize the mycoherbicide approach, law enforcement appears to have reversed its position. Semantics aside, most fungi that attack marijuana also attack hemp (McPartland 1995b, 1995c, 1997, McPartland and Cubeta 1997).
Clearly, the greatest concern surrounding biological control is host specificity. Consider Pseudoperonospora cannabina, a marijuana pathogen promoted by biocontrol researchers (Zabrin 1981, McCain and Noviello 1985). P. cannabina may be identical to Pseudoperonospora humuli, a pathogen of hemp and hop (Hoerner 1940). McPartland (1995d) investigated several fungi that were originally described as specific pathogens of Cannabis, but under closer scrutiny, turned out to be misidentifications of widespread pathogens that attack many hosts (for example, "Pleosphaerulina cannabina" turned out to be Leptosphaerulina trifolii, "Stemphylium cannabinum" = Stemphylium botryosum, "Sclerotinia kauffmanniana" = Sclerotinia sclerotiorum).
Genetic engineering
Wishing to improve host specificity and toxicity of fungal pathogens, researchers are now turning to genetic engineering (Brooker and Bruckart 1996). The use of transgenic organisms, however, elicits a new set of concerns (Levin and Israeli 1996). These are concerns that resulted in the Asilomar moratorium on genetic engineering of human pathogens.
Genetic engineers have recently been investigating a coca pathogen, Fusarium oxysporum f. sp. erythroxli (Sands et al. 1997, Nelson et al. 1997). F. oxysporum f. sp. erythroxli was selected for coca eradication because it caused natural epidemics in Peru and on the former Coca-cola plantation on Kauai, where "containment of the fungus proved challenging" (Sands et al. 1997). Fusarium oxysporum is well known to bioengineers, and previous researchers successfully inserted toxin genes into the species (Kistler 1991). Nevertheless, Gabriel (1991) considered it "unwise" to clone a toxin gene into a necrotrophic pathogen (such as F. oxysporum). He argued that such a pathogen might gain unexpected fitness and radically expand its host range, "a potentially dangerous experiment." Fusarium species can produce a variety of toxic metabolites known as trichothecenes, which gained some notoriety for their reputed use in biological warfare ("yellow rain"). F. oxysporum is known to cause systemic infections in humans (Rippon 1988).
Genetically transformed fungi have unstable genotypes, making mutations more likely. Experiments have shown F. oxysporum spontaneously mutates its transgenic DNA (Kistler 1991). Furthermore, F. oxysporum utilizes parasexual coupling, and at least 5% of its genome consists of transposons, or moveable pieces of DNA (Kistler 1997). Parasexuality and active transposable elements would facilitate the transfer of recombinant DNA to native fungi, potentially creating new strains of virulent pathogens. The wheat pathogen Puccinia graminis, for instance, hybridizes with other fungi on wild grasses, giving rise to offspring with increased virulence (Luig and Watson 1972, Burdon et al. 1981). This fact is not cited by proponents of biocontrol with rust fungi (Cook et al. 1996).
"Gene flow" has been more thoroughly studied in plants than fungi. Levin and Israeli (1996) documented five examples of spontaneous gene flow from crops to native plants, which resulted in new or worse weeds. The introgression of engineered genes from transgenic crops to related weed species has been demonstrated (Brown & Brown 1996), and may arise after just 2 generations of hybridization and backcrossing (Mikkelsen et al. 1996).
Currently, testing for gene flow is not standard procedure during the evaluation of transgenic organisms. This could be accomplished by crossing engineered fungi with related fungi (particularly if the fungi reproduce sexually, and especially if they are heterothallic fungi). Several generations of crossed hybrids are evaluated in serial host studies. Testing for gene flow is especially imperative for biocontrols which have been genetically manipulated to resist fungicides. Researchers have transformed Colletotrichum gloesporioides f. sp. aeschynomene (Collego®) with a gene for fungicide resistance (Brooker and Bruckart 1996). Imagine if this fungicide-resistant gene introgressed into Histoplasmosis capsulati or other human pathogens commonly found in agricultural areas!
The species question, round two
Another Fusarium species, F. oxysporum f. sp. cannabis (Fig. 2) is the primary candidate to kill marijuana (Hildebrand and McCain 1978, Noviello et al. 1990) and feral hemp in the American Midwest (Shay 1975). Researchers promote F. oxysporum as a marijuana mycoherbicide because they claim that hop, (Humulus lupulus), is not susceptible to fusarium wilt (McCain and Noviello 1985). However, they overlooked "Hops wilt" caused by F. oxysporum in Australia (Sampson and Walker 1982).
F. oxysporum f. sp. cannabis was originally isolated from hemp cultivars in Italy, by researchers who believed "...the wilt disease and its pathogen have not been previously described" (Noviello and Snyder 1962). In fact, these researchers missed many previous descriptions of this wilt disease (Dobrozrakova et al., 1956, Rataj 1957, Ceapoiu 1958, Czyzewska and Zarzycka 1961, Barloy and Pelhate 1962, Serzane 1962). All previous descriptions attributed hemp wilt disease to Fusarium oxysporum f. sp. vasinfectum. This fungus is morphologically identical to F. oxysporum f. sp. cannabis, but has a very broad host range (e.g., cotton, mung beans, pigeon peas, rubber trees, alfalfa, soybeans, coffee, tobacco and many other plants).
McPartland (1995a) proposed that F. oxysporum f. sp. cannabis may be a misidentified pathotype of F. oxysporum f. sp. vasinfectum. Similarly, the fungus causing tobacco wilt, originally named F. oxysporum f. sp. nicotianae, proved to be a race of F. oxysporum f. sp. vasinfectum (Armstrong and Armstrong 1975). According to Kistler et al. (1998), F. oxysporum f. sp. vasinfectum consists of at least 10 vegetative compatibility groups (VCGs). Comparing F. oxysporum f. sp. cannabis with the genotype of F. oxysporum f. sp. vasinfectum can be accomplished with VCG studies using nit mutants.
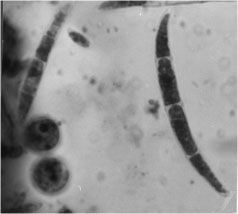 Conflicting interestsFigure 2. Microscopic spores of Fusarium oxysporum, a potential mycoherbicide of Cannabis.
Conflicting interestsFigure 2. Microscopic spores of Fusarium oxysporum, a potential mycoherbicide of Cannabis.
U.S. regulations have prevented the testing of bioengineered fungi in the field (Brooker and Bruckart 1996). But regulatory oversight is lacking in Peru and Colombia (Levin and Israeli 1996). Exigencies generated by the drug war metaphor could dangerously rush these fungi into deployment.
Moreover, saboteurs or irresponsible scientists could breach regulatory barriers, as occurred in Montana where several bioengineered organisms were illegally released around 1987 (Roberts 1987). In Australia, saboteurs illegally introduced the fungus Phragmidium violaceum to control European blackberry (Rubus fruticosus). Weedy R. fruticosus was spreading across pastures and impeding Australian cattle ranchers. The government had previously rejected ranchers’ requests to import P. violaceum, because of economic objections from commercial blackberry growers and beekeepers. Wind-borne spores of illegally introduced P. violaceum dispersed rapidly across the continent, and the fungus now infests at least four Rubus species (Watson 1991).
The Australian debacle illustrates how biocontrol may impact competing interests. The first U.S. drug czar, Carlton Turner, recognized that target plants may be considered noxious weeds by one group, and valuable crops by another group (Turner 1985). St. John’s wort (Hypericum perforatum) is an excellent example. H. perforatum was previously branded a noxious weed. But now it has become the second-best-selling herbal medicine in the U.S. — $121 million dollars of H. perforatum was sold last year, and producers are predicting a severe shortage of this raw material (Brevoort 1998).
Consultants to the European and Canadian hemp industry face a dilemma. Ecologists endorse classical (non-engineered) biocontrol organisms as potential replacements of chemical pesticides (McPartland 1984, Doctor 1986). Physicians praise the safety of biocontrols over paraquat and other synthetic herbicides (McPartland and Pruitt 1997). Nearly 20 years ago, these reasons guided the decision to search for classical biocontrols against marijuana (McPartland 1983). But times have changed. Hemp cultivation has resurged in western Europe, the former USSR, and China. Last year the Canadian government allowed farmers to grow hemp for the first time in 50 years — 251 farmers successfully harvested 5,930 acres (Cauchon 1998). Have our neighbors to the north been explicitly informed of the "Western Hemisphere Drug Elimination Act" spearheaded by Rep. McCollum? The development of transgenic mycoherbicides against marijuana would endanger hemp cultivation, permanently. Hemp is usually a pest- and disease-tolerant crop requiring little or no pesticide for cultivation. It has been characterized as "an environmentally friendly crop for a sustainable future" (Ranalli 1999). Hemp should not be endangered, and research involving transgenic pathogens of Cannabis should be halted. Moreover, the use of genetically engineered pathogens as a weapon in "the drug war" should be re-evaluated.
Acknowledgements
We thank David Morris and two anonymous phytopathologists for reviewing and improving our manuscript.
References
- Armstrong, G. and J. Armstrong 1975. Reflections on the wilt fusaria. Annual Review of Phytopathology 13:95-103.
- Auld, B. A. 1991. "Economic aspects of biological weed control with plant pathogens," in TeBeest, D. O. [Ed.] Microbial Control of Weeds Chapman & Hall, New York:262-273.
- Barloy, J. and J. Pelhate 1962. Premières observations phytopathologiques relatives aux cultures de chanvre en Anjou. Annales des Épiphyties 13:117-149.
- Booker, N. L. and W. Bruckart 1996. "Genetically engineered fungi in agriculture" in Levin, M. A. and E. Israeli [Eds.] Engineered Organisms in Environmental Settings CRC Press, Boca Raton, FL:149-163.
- Brevoort, P. 1998. The booming U.S. botanical market. HerbalGram 44:33-46.
- Brown, J and A. P. Brown 1996. Gene transfer between canola (Brassica napus L. and B. campestris L.) and related weed species. Annals Applied Biology 129: 513-522.
- Burdon, J. J., D. R. Marshall and N. H. Luig 1981. Isozyme analysis indicates that a virulent cereal rust pathogen is a somatic hybrid. Nature 293:565-566.
- Cauchon, D. 1998. Canadian hemp isn’t going to pot. USA Today 17(17)(7 Oct 1998):13-14.
- Ceapoiu, N. 1958. Cinepa, Studiu monografic. Editura Academiei Republicii Populare Romine. Bucharest. 652 pp.
- Cook, R. J. et al. 1996. Safety of microorganisms intended for pest and plant disease control: a framework of scientific evaluation. Biological Control 7:333-351.
- Czyzewska, S. and H. Zarzycka 1961. Ergebnisse der bodeninfektionsversuche an Linum usitatissimum, Crambe alyssinica, Cannabis sativa und Cucurbita pepo var. oleifera mit einigen Fusarium-Arten. Instytut Ochrony Roslin, Reguly, Polen. Report No. 41:15-36.
- Dobrozrakova, T. L. et al. 1956. "Cannabis sativa L." in Opredelitel’ Bolesni Rasteniî, Moscow:242-248.
- Doctor, B. 1986. Interview with John McEno. Sinsemilla Tips 6(1):33-34, 84-85.
- Fields, G. 1998. U.S. might enlist fungi in drug war. USA Today 17(28)(22 Oct 1998):1.
- Gabriel, D. W. 1981. "Parasitism, host species specificity, and gene-specific host cell death," in TeBeest, D. O. [Ed.] Microbial Control of Weeds Chapman & Hall, New York:115-131.
- Ghani, M., A. Basit and M. Anwar 1978. Final Report: Investigations on the natural enemies of marijuana, Cannabis sativa L. and opium poppy, Papaver somniferum L. Commonwealth Institute of Biological Control, Pakistan station. 26 pp. + 12 illus.
- Hildebrand, D. C. and A. M. McCain 1978. The use of various substrates for large scale production of Fusarium oxysporum f. sp. cannabis inoculum. Phytopathology 68: 1099-1101.
- Hoerner, G. R. 1940. The infection capabilities of hop downy mildew. J. Agric. Res. 61:331-334.
- Howarth, F. G. 1991. Environmental impacts of classical biological control. Annual Review Entomology 36:485-509.
- Kistler, H. C. 1991. "Genetic manipulation of plant pathogenic fungi" in TeBeest, D. O. [Ed]. Microbial control of Weeds Chapman & Hall, New York:152-170.
- Kistler, H. C. 1997. Genetic diversity in the plant-pathogenic fungus Fusarium oxysporum. Phytopathology 87:474-479.
- Kistler, H. C. et al. 1998. Systematic numbering of vegetative compatibility groups in the plant pathogenic fungus Fusarium oxysporum. Phytopathology 88:30-32.
- Lentz, P. L., B. R. Lipscomb and D. F. Farr 1975. Fungi and diseases of Erythroxylon. Phytologia 30:350-367.
- Levin, M. and E. Israeli 1996. "General overview of releases to date" in Levin, M. A. and E. Israeli [Eds.] Engineered organisms in environmental settings, CRC Press, Boca Raton, FL:13-39.
- Luig, N. H. and I. A. Watson 1972. The role of wild and cultivated grasses in the hybridization of formae speciales of Puccinia graminis. Aust. J. Biol. Sci. 25:335-42.
- Lockwood JA. 1993. Environmental issues involved in biological control of rangeland grasshoppers (Orthoptera: Acrididae) with exotic agents. Environmental Entomology 22:503-518.
- McCain, A. H. and C. Noviello 1985. Biological control of Cannabis sativa. Proceedings, 6th International Symposium on Biological Control of Weeds:635-642.
- McPartland, J. M. 1983. Fungal pathogens of Cannabis sativa in Illinois. Phytopathology 72:797.
- McPartland, J. M. 1984. Pathogenicity of Phomopsis ganjae on Cannabis sativa and the fungistatic effect of cannabinoids produced by the host. Mycopathologia 87:149-153.
- McPartland, J. M.. 1992. The Cannabis pathogen project: report of the second five-year plan. Mycological Society of America Newsletter 43(1):43.
- McPartland, J. M.. 1995a. Cannabis pathogens VIII: misidenfications appearing in the literature. Mycotaxon 53:407-416.
- McPartland, J. M. 1995b.Cannabis pathogens X: Phoma, Ascochyta and Didymella species. Mycologia 86: 870-878.
- McPartland, J. M. 1995c. Cannabis pathogens XI: Septoria spp. on Cannabis sativa, sensu strico. Sydowia 47:44-53.
- McPartland, J. M. 1995d. Cannabis pathogens XII: lumper’s row. Mycotaxon 54:273-279.
- McPartland, J. M. 1997. "Krankheiten und Schädlinge an Cannabis," in Symposium Magazin, 2nd Biorohstoff Hanf Technisch-wissenschaftliches Symposium. Nova Institut, Köln, Germany:37-38.
- McPartland, J. M. and M. A. Cubeta 1997. New species, combinations, host associations and location records of fungi associated with hemp (Cannabis sativa). Mycological Research 101:853-857.
- McPartland, J. M. and P. L. Pruitt 1997. Medical marijuana and its use by immunosuppressed individuals. Alternative Therapies in Health and Medicine 3(3):39-45.
- Mikkelsen, T. R., B. Andersen and R. B. Jørgensen 1996. The risk of crop transgene spread. Nature 380:31.
- Nelson, A. J. et al. Genetic characterization by RAPD analysis of isolates of Fusarium oxysporum f. sp. erthroxyli associated with an emerging epidemic in Peru. Phytopathology 87:1220-1225.
- Noviello, C. and W. C. Snyder 1962. Fusarium wilt of hemp. Phytopathology 52:1315-1317.
- Noviello, C. et al. 1990. Lotta biologica contro Cannabis sativa mediante l’impiego di Fusarium oxysporum f. sp. cannabis. Annali della Facolta di Scienze Agrarie della Universita degli Studi di Napoli, Portici 24:33-44.
- Ranalli, P. [Ed.] 1999. Advances in Hemp Research. Haworth Press, Binghamton, NY. 272 pp.
- Rata,j K. 1957. Skodlivi cinitele pradnych rostlin. Prameny literatury 2:1-123.
- Rippon, J. W. 1988. Medical Mycology, 3rd ed. W.B.Saunders Co., Philadelphia, PA. 797 pp.
- Roberts, L. 1987. New questions in Strobel case. Science 237:1098-8.
- Sampson, P. J. and J. Walker 1982. An annotated list of plant diseases in Tasmania. Dept. of Agriculture, Tasmania, Australia 121 pp.
- Sands, D. C., et al. 1997. Characterization of a vascular wilt of Erythroxylon coca caused by Fusarium oxysporum f. sp. erythroxyli forma specialis nova. Plant Disease 81:501-504.
- Schmitt, C. G. and B. R. Lipscomb 1975. Pathogens of elected members of the Papaveraceae — an annotated bibliography. USDA-ARS Northeastern Region Report No. 62. USDA-ARS, Beltsville, MD. 186 pp.
- Schultes, R. E., W. M. Klein, T. Plowman and T. E. Lockwood 1974. Cannabis: an example of taxonomic neglect. Bot. Mus. Leaflet. Harv. Univ. 23:337-367.
- Serzane, M. 1962. "Kanepju - Cannabis sativa L. Slimibas." in Augu Slimibas, Praktiskie Darbi. Riga Latvijas Valsts Izdevnieciba, Lativa USSR.: 366-369.
- Shay, R. 1975. Easy-gro fungus kills pot among us. The Daily Californian, March 14:3.
- Small, E. 1979. The species problem in Cannabis. Volume 2: semantics. Corpus Information Services Ltd. and Agriculture Canada. Ottawa. 156 pp.
- Small, E. and A. Cronquist 1976. A practical and natural taxonomy for Cannabis. Taxon 25:405-435.
- TeBeest, D. O. 1988. Additions to host range of Colletotrichum gloeosporiodes f. sp. aeschynomene. Plant Disease 72:16-18.
- Turner, C. E. 1985. "Conflicting interests and biological control of weeds," in Proceedings 6th International Symposium Biological Control of Weeds: 203-225 .
- Watson, A. K. 1991. "The classical approach with plant pathogens," in TeBeest, D. O. [Ed.] Microbial Control of Weeds, Chapman & Hall, New York:3-23.
- Zubrin, R. 1981. The fungus that destroys pot. War on Drugs Action Reporter: June 1981:61-62.
Editor’s NoteFor additional reading on this timely subject see; Kleiner, Kurt 1999 "Operation Eradicate" in New Scientist Sept. 11 with the accompanying editorial and Hogshire, Jim 1998 "The Drug War’s Fungal Solution?" in Covert Action Spring issue.
grünegrüsse
hans


