Light Regulation of Stomatal Movement
Plants have evolved multiple photoreceptor systems to monitor light quality, quantity and direction.
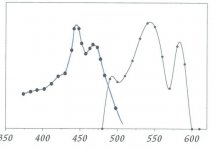
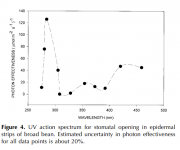
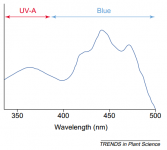
It is well known that stomatal morphogenesis is controlled by genetic as well as environmental factors and in general, an increase in light intensity results in an increase in stomatal index and a systemic signal from mature leaves determine the response.43,75–78 Light is perceived by various photoreceptors,79,80 and stomatal movements are regulated by both blue and red light.
The blue-light response of stomata appears to be strongly affected by red light, and the underlying mechanisms in the interaction between blue-light signaling and guard cell chloroplasts were recently reviewed in reference 14.
It is shown that blue light-induced stomatal opening is mediated by the blue light receptor phototropins (PHOT1 and PHOT2) and cryptochromes (CRY1 and CRY2).
81 Recent findings suggest that the light control of stomatal development is mediated through a crosstalk between the cryptochrome-phytochrome-COP1 signaling system and the mitogen-activated protein kinase signaling pathway.
82 Blue light is required for the activation of phototropins, plant-specific Ser/Thr autophosphorylating kinases, and the activated phototropins transmit the signal to the plasma membrane H+-ATPase for its activation.
80,83 Activation of the H+-ATPase is caused by the phosphorylation of a Thr residue in the C terminus with subsequent binding of a 14-3-3 protein to the Thr residue.84,85 Since phototropins are Ser/Thr protein kinases, it might be possible that phototropins directly phosphorylate the H+-ATPase. However, this has been shown not to be the case since it was demonstrated that protein phosphatase 1 (PP1), a major member of the PPP family of Ser/Thr protein phosphatases, mediates the signaling between phototropins and H+-ATPase in guard cells.86 Therefore, ABA is likely to inhibit the signaling molecule(s), including phototropins, PP1, H+-ATPase and other unidentified components.
It has also been suggested that the guard cell response to red light is in part an indirect response to red-light-driven intercellular CO2 uptake in the mesophyll.87 For example, it has been shown that chloroplast-containing guard cells in albino sections of variegated leaves do not respond to photosynthetically active radiation, but are sensitive to blue light and CO2, bringing into question a direct role of guard cell photosynthesis on red-lightmediated stomatal opening in intact leaves.35
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3258058/
Plants have evolved multiple photoreceptor systems to monitor light quality, quantity and direction.
It is well known that stomatal morphogenesis is controlled by genetic as well as environmental factors and in general, an increase in light intensity results in an increase in stomatal index and a systemic signal from mature leaves determine the response.43,75–78 Light is perceived by various photoreceptors,79,80 and stomatal movements are regulated by both blue and red light.
The blue-light response of stomata appears to be strongly affected by red light, and the underlying mechanisms in the interaction between blue-light signaling and guard cell chloroplasts were recently reviewed in reference 14.
It is shown that blue light-induced stomatal opening is mediated by the blue light receptor phototropins (PHOT1 and PHOT2) and cryptochromes (CRY1 and CRY2).
81 Recent findings suggest that the light control of stomatal development is mediated through a crosstalk between the cryptochrome-phytochrome-COP1 signaling system and the mitogen-activated protein kinase signaling pathway.
82 Blue light is required for the activation of phototropins, plant-specific Ser/Thr autophosphorylating kinases, and the activated phototropins transmit the signal to the plasma membrane H+-ATPase for its activation.
80,83 Activation of the H+-ATPase is caused by the phosphorylation of a Thr residue in the C terminus with subsequent binding of a 14-3-3 protein to the Thr residue.84,85 Since phototropins are Ser/Thr protein kinases, it might be possible that phototropins directly phosphorylate the H+-ATPase. However, this has been shown not to be the case since it was demonstrated that protein phosphatase 1 (PP1), a major member of the PPP family of Ser/Thr protein phosphatases, mediates the signaling between phototropins and H+-ATPase in guard cells.86 Therefore, ABA is likely to inhibit the signaling molecule(s), including phototropins, PP1, H+-ATPase and other unidentified components.
It has also been suggested that the guard cell response to red light is in part an indirect response to red-light-driven intercellular CO2 uptake in the mesophyll.87 For example, it has been shown that chloroplast-containing guard cells in albino sections of variegated leaves do not respond to photosynthetically active radiation, but are sensitive to blue light and CO2, bringing into question a direct role of guard cell photosynthesis on red-lightmediated stomatal opening in intact leaves.35
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3258058/

") I don't think I've read that before.
I don't think I've read that before.